Caspases are the family of cysteine aspartate-specific proteases that play a critical role in proteolytic execution, with significant roles in apoptosis, necrosis, cytokine maturation, inflammatory responses, cellular differentiation, and development. [show the full text]
Caspase 억제제/활성제 (Caspase Inhibitors/Activators)
기타 Apoptosis 억제제
| Cat.No. | 제품명 | 정보 | 제품 사용 인용 | 제품 검증 |
|---|---|---|---|---|
| S7775 | Emricasan (IDN-6556) | Emricasan (IDN-6556, PF 03491390, PF-03491390)은 강력한 비가역적 pan-caspase 억제제입니다. Emricasan은 지카 바이러스 감염 억제제입니다. |
|
|
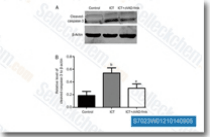
| S7023 | Z-VAD-FMK | Z-VAD-FMK (Z-VAD(OMe)-FMK)는 세포 투과성 비가역적 팬-caspase 억제제로, THP.1 및 Jurkat T-세포에서 apoptosis의 모든 특징을 차단합니다. |
|
|
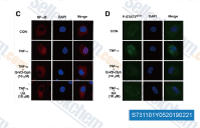
| S7311 | Q-VD-Oph | Q-VD-Oph (Quinoline-Val-Asp-Difluorophenoxymethylketone)는 카스파제 1,3,8 및 9에 대해 25 내지 400 nM 범위의 IC50을 갖는 강력한 팬-caspase 억제제입니다. Q-VD-OPh는 HIV 감염을 억제할 수 있습니다. |
|
|
| S7312 | Z-DEVD-FMK | Z-DEVD-FMK (Caspase-3 Inhibitor)는 특이적이고 비가역적인 Caspase-3 억제제이며, Caspase-6, Caspase-7, Caspase-8 및 Caspase-10에도 강력한 억제 효과를 보입니다. |
|

|
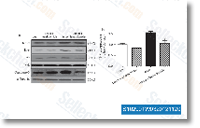
| S1029 | CC-5013 (Lenalidomide) | Lenalidomide는 PBMC에서 IC50이 13 nM인 TNF-α 분비 억제제입니다. Lenalidomide (CC-5013)는 ubiquitin E3 ligase cereblon (CRBN)의 리간드이며, CRBN-CRL4 ubiquitin ligase에 의해 두 가지 림프계 전사 인자인 IKZF1 및 IKZF3의 선택적 유비퀴틴화 및 분해를 유발합니다. Lenalidomide는 cleaved caspase-3 발현을 촉진하고 VEGF 발현을 억제하며 apoptosis를 유도합니다. |
|
|
| S9042 | Wedelolactone | 약용 식물에서 유래한 천연 화합물인 Wedelolactone은 IκBα의 인산화 및 분해를 매개하여 NF-κB 활성화에 중요한 IKK의 억제제입니다. 이 화합물은 또한 caspase-11의 억제제입니다. |
|
|
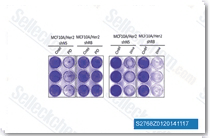
| S2768 | Dinaciclib (SCH 727965) | Dinaciclib은 세포를 사용하지 않는 분석에서 CDK2, CDK5, CDK1 및 CDK9에 대한 새로운 강력한 CDK 억제제로, IC50은 각각 1 nM, 1 nM, 3 nM 및 4 nM입니다. 또한 티미딘(dThd) DNA 통합을 차단합니다. Dinaciclib은 Caspase 8 및 9 활성화를 통해 세포자멸사를 유도합니다. 3상. |
|
|
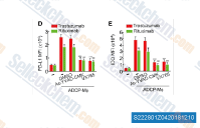
| S2228 | Belnacasan (VX-765) | VX-765 (Belnacasan)는 세포 유리 분석에서 Ki가 0.8 nM인 caspase-1의 강력하고 선택적인 억제제이며, 2단계에 도달했습니다. |
|
|
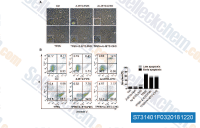
| S7314 | Z-IETD-FMK | Z-IETD-FMK (Caspase-8 Inhibitor, Z-IE(OMe)TD(OMe)-FMK)는 특정 Caspase-8 억제제입니다. Z-IETD-FMK는 또한 그랜자임 B 억제제이기도 합니다. |
|
|
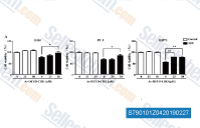
| S7901 | Ac-DEVD-CHO | Ac-DEVD-CHO (Caspase-3 Inhibitor I, N-Ac-Asp-Glu-Val-Asp-CHO)는 그룹 II 카스파제에 대한 강력한 알데히드 억제제로, Caspase-3 및 Caspase-7에 대해 각각 0.2 nM 및 0.3 nM의 Ki 값을 가집니다. Caspase-2에 대한 약한 억제력을 보입니다. |
|
|
Caspases can be traditionally divided into two groups based on their sequence homology and function. Caspase-1, -4 and -5 belong to Group I (inflammatory) caspases (caspase-1-related subfamily) that are involved in cytokine maturation and the innate immunity. The Group II caspases (caspase-3-related subgroup) are involved in the regulation of apoptosis, which are further divided into two types: initiators (apical caspases) that includes caspase-2, -8, -9, and -10, and effectors (executing caspases) such as caspase-3, -6 and 7. These caspases have distinct substrate cleavage specificities. To date, over 600 substrates for the cell death-related caspases have been identified. Effector caspases are constitutively produced in cells as dimmers, and the proteolytic processing into cleaved caspases by an initiator enzyme is required to trigger their activity. Being active, effector caspases target a wide spectrum of cellular proteins with the ultimate effect of causing cell death. In contrast to effector caspases, initiator caspases are translated as monomeric zymogens. Formation of multicomponent complexes triggers initiator caspase dimerization sufficient for their activation. Recently, a novel alternative perspective is proposed that mammalian caspases are activated, not to kill, but to extinguish the pro-inflammatory properties of dying cells. This perspective unifies the mammalian caspase family as either positive or negative regulators of inflammation. [1][2]
Caspase-1 subfamily members (caspase-1, -4, and -5) have been implicated as regulators of inflammation through processing and activating two related cytokines, IL-1β and IL-18. The initiator caspases are activated by upstream molecules through protein-protein interaction domains known as caspase recruitment domain (CARD) and death effector domain (DED). The death-inducing signaling complex (DISC), the Apaf-1 apoptosome and the p53-induced protein with a death domain (PIDD) are protein assembly platforms that can recruit caspase-8/-10, -9 and -2, respectively, confirming the essential roles of caspases in both the extrinsic receptor-mediated and intrinsic mitochondrial apoptosis pathways. Caspases are regulated at a post-translational level by inhibitors of apoptosis and by dominant negative isoforms. The proteolytic activity of mature caspase-9 and -3 are subdued by the inhibitor of apoptosis proteins (IAPs). In turn, IAPs are inactivated and caspase activity restored by proteins, such as SMAC/Diablo or HtrA2/Omi, which are released from the mitochondria. The cellular FLICE inhibitory protein (c-FLIP) is a catalytically inactive homologue of caspase-8 and -10, which can prevent their activation by obstructing binding sites on the DISC. Hematopoietic stem cells express a smaller variant, caspase-8L, which acts as a dominant negative when recruited to DISC after CD95 triggering, thereby disrupting the link between CD95 and the caspase cascade. The pro-domain-only polypeptides of caspase-10 have been reported to be pro-apoptotic in some experimental systems, but appeared to be antiapoptotic and capable of inducing NF-κB activity in others. Moreover, either oncogenes (Myc) or tumor suppressors (p53) are able to adjust the intrinsic or the extrinsic caspase cascade involved in the complex signaling system controlling apoptosis. [1][2]
Cells do not necessarily undergo caspase-independent cell death in the absence of active caspases, but may instead survive insult and even promote clonogenic tumor growth. The loss of even one caspase-2 allele results in increased cell proliferation as well as accelerated tumorogenesis, and the loss of caspase-2 expression has been observed in gastric cancer. In a screen of primary breast tumor samples, approximately 75% of the tumors as well as morphologically normal peritumoral tissue samples lack caspase-3 transcripts and caspase-3 protein expression. Reduction of caspase-8 expression has been found in pediatric tumors, and colorectal, gastric, or hepatocellular cancers, as well as in clinical glioma samples. Downregulation of the mitochondrial IAP antagonist Smac/Diablo is associated with renal cell carcinomas, and overexpression of Survivin, another IAP, has been observed in most transformed cell lines and cancers. Caspase-3 and caspase-6 cleavage of Tau protein leads to neurofibrillary tangle formation during Alzheimer’s disease pathogenesis. Moreover, accumulated caspase-6 cleavage-mediated huntingtin fragments represent an early pathological change in the brains of Huntington’s disease patients. Conversely, an increase in cell death is associated with heart disease, stroke, neurodegenerative disorders and liver disease. Additionally, abnormal fluctuations in cytokine levels as a result of the inflammatory response have been implicated in several diseases, including osteoarthritis (OA) and rheumatoid arthritis (RA), gout, inflammatory bowel disorders, sepsis, and inflammatory skin diseases. Thus, the disturbances in the regulation of caspase activation are central for the avoidance of cell death, and have been implicated in the pathogenesis of many disorders including stroke, Alzheimer's disease, myocardial infarction, cancer, and inflammatory disease, which stimulates interest in caspases as potential therapeutic targets. Pralnacasan and VX-765 are reversible caspase-1 inhibitors that are developed for the treatment of a variety of inflammatory disorders disorders, including RA and OA. There is a large pool of inactive procaspase-3 in some cancer cells compared with normal cells, thus, targeting procaspase-3 directly with small molecule activators such as PAC-1 and 1541, rather than targeting upstream regulators of apoptosis could lead to a more effective and direct therapy as caspase-3 is the terminal protease in the apoptotic cascade. [1][3]
제품은 연구용으로만 사용됩니다. 인체에는 사용하지 마십시오. 환자에게 판매하지 않습니다.
©Copyright 2013 Selleck Chemicals. All Rights Reserved.